Tesina de Licenciatura en Química. Junio de 2010. Tutor: Prof. Dr. Andrés González Ritzel. Colaboración: Prof. Dr. Enrique Castiglioni.
Laboratorio de Ecología Química. Facultad de Química. Universidad de la República. Montevideo, Uruguay.
Resumen
En éste trabajo se realizó la colecta de compuestos orgánicos volátiles (VOCs) de soja a campo, y el análisis de los mismos en el laboratorio para determinar los perfiles de emisión en los diferentes estadios de desarrollo fenológico. Además se realizó una evaluación de los mismos frente a individuos de P. guildinii (principal plaga de la soja en Uruguay) por ensayos comportamentales y estudios electrofisiológicos (GC-EAD) en la búsqueda de posibles kairomonas utilizadas por los insectos para ubicación de su fuente alimenticia.
Laboratorio de Ecología Química. Facultad de Química. Universidad de la República. Montevideo, Uruguay.
Resumen
En éste trabajo se realizó la colecta de compuestos orgánicos volátiles (VOCs) de soja a campo, y el análisis de los mismos en el laboratorio para determinar los perfiles de emisión en los diferentes estadios de desarrollo fenológico. Además se realizó una evaluación de los mismos frente a individuos de P. guildinii (principal plaga de la soja en Uruguay) por ensayos comportamentales y estudios electrofisiológicos (GC-EAD) en la búsqueda de posibles kairomonas utilizadas por los insectos para ubicación de su fuente alimenticia.
Introducción
Glycine max, la soja.
La soja, Glycine max (L.) Merrill es una planta anual erecta de 50-90 cm. de altura perteneciente a la familia Fabaceae, subfamilia Faboideae. Sus hojas son compuestas trifoliadas, salvo las basales que son unifoliadas y se caen antes de que la planta llegue a su estado de madurez fisiológica. Sus flores son pequeñas de alrededor de 8 mm de longitud de corola blanca o violácea y los frutos son legumbres o vainas péndulas (figura 1).

Figura
1: Glycine max; planta en estadio reproductivo y semillas.
La soja es originaria de Asia Oriental, donde es
cultivada desde la antigüedad, y desde allí se difundió a los países
occidentales basado en su gran potencial alimenticio. Sus semillas son ricas en
proteínas y aceites, lo que la hace sustituta de muchos productos de origen
animal como la carne. También es de gran utilidad para la producción de
biocombustibles y plásticos. Presenta un importante contenido de isoflavonoides
que previenen el cáncer de mama, entre otros.
Existen varios cientos de variedades de soja que
difieren por el color y las formas de las semillas, así como por las
características morfológicas de las plantas. Se caracteriza por ser una especie
muy sensible a las variaciones microclimáticas, por lo cual requiere un
cuidadoso trabajo de selección de variedades antes de encontrar la de mayor
rendimiento para una región dada. Como todas las leguminosas, es
una excelente especie para utilización en esquemas de rotación de cultivos,
debido a la capacidad de fijar nitrógeno de las bacterias simbióticas del
género Rhizobium que habitan sus
nódulos radicales.
Existe un convención entre especialistas para la
descripción de los estadios de desarrollo fenológico de la soja donde los
estadios vegetativo y reproductivo son descriptos separadamente.
Los estadios vegetativos son determinados por conteo del número de nudos del
tallo principal, comenzando por el nudo unifoliado. Los estados reproductivos R1
y R2 corresponden a la floración, R3 y R4 al
desarrollo de las vainas (llenado), R5 y R6 responden al
desarrollo de las semillas, y, R7 y R8 a la madurez
fisiológica (ver figura 2). Este sistema es aplicado tanto para plantas
aisladas como para cultivos. La cantidad de estadios vegetativos
varía según la variedad de soja que se esté considerando y las condiciones
ambientales reinantes. Por ejemplo Fehr et. al observaron el comienzo de la
floración desde los 4 hasta 18 nudos en el tallo principal. Para éste sistema
descriptivo se consideran los estadios reproductivos basados en el desarrollo
de la porción superior del tallo, independizando del genotipo y ambiente de la
planta. Así R1 designa la existencia de una flor en algún nudo
(generalmente el superior) y R2 designa la presencia de floración en
el nudo inmediatamente por debajo del superior .

Figura
2: Estadios de desarrollo fenológico de la soja, según
Fehr et. al.
Nota:
en el diagrama no se presentan estados vegetativos superiores a V3,
pero su existencia depende en gran medida de la variedad de soja plantada y las
condiciones ambientales (ver texto).
Expansión del cultivo de soja. Situación en Uruguay.
La expansión
masiva de la soja se dio a principios de
la década del setenta, sobre todo en el Medio Oeste estadounidense, y a partir
de los noventa alcanzó gran auge en el Cono Sur. Hoy en día, Brasil y Argentina
se han convertido en el segundo y tercer productor mundial respectivamente
(luego de EEUU). Uruguay no ha escapado a ésta realidad, y, actualmente la soja
es el cultivo con la mayor superficie implantada en el país. La misma
evolucionó desde las 9.000 hectáreas en la temporada 1998/1999 a 400.000
hectáreas en la temporada 2007/2008 (habiendo predicciones de llegar a las
600.000 has en la zafra venidera) con una evolución concomitante en las exportaciones. La soja se cultiva principalmente en el litoral oeste del país
(Fuente MGAP, ver figura 3) y la producción anual alcanza las 800.000 toneladas.
En las últimas décadas se ha producido, a través de
técnicas de Ingeniería Genética, soja resistente a glifosato (herbicida), lo cual
ha aumentado la producción al disminuir la incidencia negativa de las malezas.
Ello ha provocado controversia sobre los efectos de dicho cultivo en el
ambiente y sus efectos en la salud pública. En el país la introducción de dicha
variedad fue a partir de la segunda mitad de los noventa, y en la actualidad,
casi la totalidad de la de soja plantada es transgénica (Soja RR; Round-up
Ready, con la incorporación de un gen de Agrobacterium
sp.).


Figura 3: Superficie sembrada de soja en
la temporada 2004-2005.
Los impactos ambientales más fuertes constatados se
relacionan con la masiva aplicación de agroquímicos al cultivo. El empleo de
aquellos ha aumentado un 400 % en los últimos 5 años. El
paquete tecnológico de la soja transgénica, incluye además del glifosato,
funguicidas e insecticidas altamente contaminantes, tales como el endosulfán,
clorpirifos, y cipermetrina entre otros. Los mismos han demostrado ser muy
nocivos para la vida terrestre y acuática, con un alto potencial de
bioacumulación. Su persistencia en el ambiente hace que aumenten
sus impactos negativos, tanto en la salud humana como en la flora y fauna,
provocando desequilibrios en los ecosistemas. En éste sentido se ha constatado
grave afectación a los enemigos naturales y resurgencia de plagas. En el caso
del endosulfán (organoclorado), existe una campaña a nivel mundial para su
eliminación y con respecto al clorpirifos, su uso ya está estrictamente
restringido en algunos países de América Latina.
Piezodorus guildinii, la chinche de las leguminosas.
Los
insecticidas son empleados para el combate de plagas, siendo las principales
que afectan al cultivo las orugas (lepidópteros) y chinches (hemípteros). Los
lepidópteros de mayor significación
económica son principalmente dos: Epinotia aporema y Anticarsia gemattalis;
mientras que la chinche más importante es Piezodorus
guildinii
(figura 4).
P. guildinii
posee, como todos los hemípteros, un aparato bucal pico-suctor que le permite penetrar
los tejidos vegetales para alimentarse (daños primarios), posibilitando la
introducción de bacterias, virus y hongos que pueden causar enfermedades a la
planta (daños secundarios). Se ha reportado adicionalmente un daño conocido
como “soja loca”, que consiste en la retención foliar de la planta al alcanzar
la madurez, lo que dificulta la recolección mecánica de las semillas.
Esta especie se encuentra en los sistemas de producción durante todo el año,
oscilando entre el cultivo de soja y las leguminosas forrajeras.
Debido a su alto nivel poblacional generalmente presente en los
agroecosistemas y su ataque directo a
las semillas; afecta el rendimiento y la calidad de las mismas. El ataque se da
en estados de desarrollo avanzado, momento en que la planta no está en
condiciones de compensar los efectos de dicho ataque, convirtiéndola en la
plaga más importante del cultivo.
Al
poner en peligro el éxito económico del negocio obliga a un estricto control en
base a la utilización de insecticidas no selectivos, en general endosulfán. Este insecticida se
aplica tanto puro como en mezclas con thiametoxán, cipermetrina u otros piretroides
equivalentes, con el objetivo de potenciar su efecto. Se debe
resaltar que hasta el momento no existen insecticidas selectivos para el
control de ésta plaga, que puedan oficiar como alternativa al uso de
endosulfán. Según Castiglioni P. guildinii ha comenzado a
desarrollar importantes niveles de resistencia a algunos de los productos
utilizados para su control, entre ellos endosulfán.
Las
aplicaciones de pesticidas en general son más frecuentes a partir de la
floración y se intensifica hasta alcanzar la madurez, período en el cuál es más
probable y dañino el ataque de la chinche.
El
control de plagas alternativo, por rotación de cultivos-pasturas o por enemigos
naturales no es empleado en la actualidad.

Figura
4:
Piezodorus guildinii; adulto y ninfas de cuarto estadío, ambas consideradas
plagas importantes del cultivo de soja.
Todos
los inconvenientes numerados permiten visualizar la no sostenibilidad de los
agroecosistemas sojeros a largo plazo. Por ello son necesarias nuevas
alternativas de manejo agronómico, y dentro de ellas medidas de control de
plagas más amigables con el medio ambiente.
Comunicación química:
los insectos y su relación con el medio.
Entre los animales,
los insectos constituyen el taxón que más frecuentemente utiliza sustancias
químicas (semioquímicos) en procesos de comunicación y percepción del medio.
Los semioquímicos involucrados en comunicación intraespecífica se conocen como
feromonas, mientras que los que median comunicación interespecífica son
conocidos como aleloquímicos. Éstos suelen dividirse para su estudio en
alomonas y kairomonas: las primeras denotan señales químicas adaptativamente
favorables para el emisor de la señal, mientras que las segundas benefician al
receptor de la señal. En las relaciones planta-insecto, las kairomonas más
conocidas son aquellas que contribuyen a la localización por parte de los
insectos fitófagos de sus plantas alimenticias.
Dado los progresos alcanzados en los últimos años en
la comprensión de los mecanismos de comunicación química entre plantas, y cómo
las plantas se defienden de sus agresores herbívoros o patógenos; el uso de
semioquímicos cómo herramienta en manejo integrado de plagas (MIP) está llamado
a ser una alternativa viable al empleo convencional de pesticidas de síntesis.
Resultados alentadores se han obtenido de la manipulación comportamental de
insectos en estrategias de push-pull, a
través del uso de feromonas para monitoreo poblacional o confusión sexual, y a
través del empleo de elicitores químicos que incrementen el nivel de defensas
directas e indirectas de la planta.
Los VOCs, moléculas señal en la naturaleza.
Los compuestos orgánicos volátiles (VOCs) son
compuestos químicos liberados a la atmósfera (alta presión de vapor) por
fuentes biogénicas o no biogénicas. Se caracterizan por ser
compuestos de bajo peso molecular (menor a 500 dalton), en su mayor parte
lipofílicos, que responden a una gran diversidad de grupos químicos.
En el caso de las fuentes biogénicas, especialmente en plantas, son
característicos los derivados isoprenoides (hemiterpenos, monoterpenos y sesquiterpenos), alcanos, alquenos, cetonas,
aldehídos, ésteres, éteres, ácidos e hidrocarburos aromáticos. Los terpenos son
los compuestos más prominentes en la mayoría de los casos. La emisión de VOCs
en plantas no es homogénea y varía apreciablemente tanto en cantidad como
identidad de los compuestos. Ello depende de factores propios del organismo
como el estado de desarrollo y genotipo, y de factores ambientales que provocan
stress tales como la contaminación, la presencia de organismos detrimentales
(fitófagos, patógenos), la duración del fotoperíodo y la temperatura.
La antedicha dependencia es característica en la biosíntesis de metabolitos
secundarios.
El entendimiento del rol de los VOCs en las interacciones
planta-insecto es menor que para los compuestos no volátiles debido las
dificultad en su aislamiento, identificación y cuantificación (se encuentran en
proporción de ppm o ppb respecto al peso de la planta) y por su naturaleza
disipativa. Ello influye en última instancia en el entendimiento del
metabolismo y la fisiología vegetal. Varios métodos han sido
empleados para aislar VOCs, entre ellos destilación por arrastre con vapor,
destilación a vacío, extracción con solventes, condensación en frío y trampas
de adsorbentes. Esta última opción es la más adecuada y
representativa de las condiciones de emisión in vivo, minimizando contaminantes, y fue la
elegida para éste trabajo.
Los VOCs de plantas han sido reportados cómo
atrayentes de insectos a sus plantas alimenticias (cumpliendo la función de kairomonas)
pero quizá su rol más importante sea como repelentes y deterrentes de fitófagos
que hagan a una planta no alimenticia (alomonas). El
comportamiento de los insectos es influido tanto por diferencias cualitativas
cómo cuantitativas en la pluma de volátiles.
Liu et al han demostrado que las distintas
variedades de soja emiten diferentes perfiles de volátiles que pueden influir
en la aceptabilidad por parte de un insecto fitófago. Las especies cultivadas
parecen ser más apetecibles a los mismos que las especies silvestres.
En la bibliografía se hace una distinción entre los
VOCs emitidos por la planta en condiciones normales y los VOCs emitidos bajo
herviboría. Éstos últimos son conocidos como HIPVs (herbivore-induced plant volátiles,
en inglés) los que generalmente tienen un rol relevante en interacciones
tritróficas en la atracción de depredadores o parasitoides del herbívoro
(incluso en el caso de volátiles inducidos por pentatómidos; figura 5).

A la atracción de dichos enemigos naturales por parte de volátiles de la planta se le llama defensa indirecta, y las plantas suelen recurrir a ellas cuando es bajo el nivel de defensas directas (constitutivas o inducidas por la misma planta). Los compuestos que generalmente median la liberación de defensas inducidas son salicilato de metilo (MeSA) y ácido jasmónico. Los mismos son liberados por la planta luego de una cascada de señalización iniciada tras el inicio de la alimentación u oviposición. La sola presencia de dichos compuestos activa los mecanismos de defensa de la planta y la atracción de los parasitoides, no necesitándose la presencia del insecto perjudicial.
El MeSA también está involucrado en interacciones tritróficas en la naturaleza. Zhu et. al demostraron que las plantas de soja altamente atacadas por Aphis glyicine (Homoptera:Aphidae) emiten en alta proporción MeSA, lo que provoca una respuesta en GC-EAD de su principal depredador, Coccinella setempuntacta (Coleoptera:Coccinellidae).
Figura 5: Atracción de una avispa parasitoide por parte de los volátiles de hoja verde (GLVs) liberados por la herbivoría de una oruga.
A la atracción de dichos enemigos naturales por parte de volátiles de la planta se le llama defensa indirecta, y las plantas suelen recurrir a ellas cuando es bajo el nivel de defensas directas (constitutivas o inducidas por la misma planta). Los compuestos que generalmente median la liberación de defensas inducidas son salicilato de metilo (MeSA) y ácido jasmónico. Los mismos son liberados por la planta luego de una cascada de señalización iniciada tras el inicio de la alimentación u oviposición. La sola presencia de dichos compuestos activa los mecanismos de defensa de la planta y la atracción de los parasitoides, no necesitándose la presencia del insecto perjudicial.
El MeSA también está involucrado en interacciones tritróficas en la naturaleza. Zhu et. al demostraron que las plantas de soja altamente atacadas por Aphis glyicine (Homoptera:Aphidae) emiten en alta proporción MeSA, lo que provoca una respuesta en GC-EAD de su principal depredador, Coccinella setempuntacta (Coleoptera:Coccinellidae).
Está bien caracterizada la emisión por parte de las
plantas de volátiles de hoja verde (green leaf volatiles, GLV, en inglés), los
cuales son alcoholes, cetonas, aldehídos y ésteres alifáticos de hasta 10
átomos de carbono, emitidos por todas las plantas en condiciones in vivo. Muchos de ellos han demostrado
tener funciones importantes en la naturaleza; Gardner et al reportan la inhibición de germinación de semillas y del crecimiento de plántulas de soja
por parte del hexanal, E-2-hexenal y E-2-nonenal.
Materiales y Métodos
Materiales y Métodos
Colecta de VOCs a campo:
Ésta parte del
trabajo se realizó en el período enero-abril de 2009, en una campo de soja en
la Estación Experimental “Mario Cassinoni” (EEMAC) de Facultad de Agronomía
(UdelaR), Paysandú.
Para ello se muestreó
en cinco estadios de desarrollo fenológico del cultivo de soja: vegetativo (V4-V5
y V8-V9), floración (R2), llenado de vaina (R5)
y madurez fisiológica (R8). En todos los casos la colecta se realizó
de manera semejante.
El dispositivo
empleado para la colecta a campo se muestra en la figura 6.

Figura 6: Sistema de colecta para
volátiles a campo, aquí ejemplificado para el muestreo en estadios V8-V9.
La idea básica fue
captar los volátiles (VOCs) in vivo
de las plantas de soja. El follaje de las
plantas de soja fue embolsado a cierta altura del suelo (por sobre las hojas
unifoliadas) de manera de no captar volátiles del mismo; formando una cámara
abierta por su extremo inferior. Las bolsas empleadas fueron de poliestireno
(Embale Bem, 27 x 41 cm). En el extremo superior de la misma se adosó una
columna de vidrio (tipo pipeta Paster; 15.7 cm l. x 0.6 cm d. i.) empacada con
0.1 g de Super Q® entre dos porciones de lana de vidrio.
Simultáneamente se preparó un blanco con una bolsa vacía. Las salidas de ambas
columnas fueron unidas a un conector en forma de “T” en cuyo extremo se colocó
una bomba de vacío portátil (Apex 1.04; Casella Cel). Así la bomba arrastró la
atmósfera circundante a las plantas y los compuestos volátiles fueron recogidos
en el lecho de adsorbente. Las condiciones de muestreo fueron las siguientes:
flujo de aire: 1.60 L/min; programa de muestreo: 20 horas; sistemas de colecta
por duplicado. La temperatura fue variable en virtud de la variación climática
diaria. Las bombas fueron colocadas a cierta altura del suelo y protegidas
mediante malla sombra dado que al ser expuestas al sol directamente, las
condiciones de funcionamiento pueden variar. Las bolsas también fueron
parcialmente protegidas del calor mediante malla sombra para evitar el
marchitamiento de las plantas muestreadas. Con ello también se mitiga en parte
la descomposición, como consecuencia del calor, del material constituyente de
las bolsas.
Extracción de VOCs:
Para la extracción de
los VOCs se procedió a la elución de las columnas provenientes de los muestreos
(tanto de blanco como de soja) con 2 ml de n- hexano (95 % de pureza,
Mallinckrodt Chemicals), adicionándose 100 µl de ntridecano (0.05408 mg/ml)
cómo estándar interno. A dicho extracto se lo concentró bajo corriente gaseosa
de N2 hasta 100 µl, y se lo almacenó a -18 ºC hasta su posterior
análisis químico y comportamental.
Análisis por cromatografía gaseosa (GC):
Para ello se empleó
un cromatógrafo Hewlett-Packard 5890 serie II con las siguientes condiciones:
gas carrier: H2; T inyector: 220 ºC; T detector (FID): 250 ºC;
método: 40 ºC (4 min)- 10 ºC/min- 240 ºC (5 min); Inyección Splitless, Columna
Elite 5 (5% difenil- 95% dimetil polisiloxano; 30 m x 0.25 mm d.i. x 0.25 μm).
El cromatógrafo se
conectó a una PC equipada con el Software HP Chem Station, donde pudo
visualizarse los cromatogramas y ajustar las condiciones de funcionamiento del
equipo.
La determinación de
picos pertenecientes a extractos de soja fue realizada por comparación entre
los cromatogramas de los mismos y los correspondientes de los blancos.
Se calcularon los
índices de retención (IR) de los componentes de los extractos de VOCs de soja
por inyección de 1 µl del extracto bajo las mismas condiciones cromatográficas
que en el caso anterior, en paralelo a 0.5 µl de solución de Kovats (C8,
C10, C12, C14, C16, C18,
C19, C20, C22, C26, C32; 100
ppm).
Análisis por GC acoplado a espectrometría de masas
(GC-MS):
Los extractos de
todos los estadios fueron inyectados en modalidad splitless por duplicado (1
µl) en un GC-MS Shimadzu QP2010 Plus. La columna empleada fue OPTIMA- 5-MS (30
m x 0.25 mm x 0.25 μm) con He como gas portador. La fuente de ionización fue de
impacto electrónico (IE) a 70-eV sometida a 200 ºC y con la línea de
transferencia a 300 ºC. La cuantificación fue realizada por comparación de
áreas con el estándar interno de n-tridecano. Adicionalmente se inyectó 0.5 µl
de la solución de Kovats para un segundo cálculo de índices de retención (IR).
La identificación de
los compuestos fue mediante comparación de sus IR y espectros de masas con la
datos de la base de datos SHIM 2205 (15).Como criterio de identificación
se tomó un porcentaje de similitud de espectros mayor al 85%, y una diferencia
de índices de retención no mayor a 10 unidades. Identificación tentativa se le
llamó a aquella en la que sólo se pudo contar con el porcentaje de similitud
del espectro de masas, tomándose en éste caso como criterio un 90% de similitud
en los mismos. Aquellos compuestos que
se encontraron en los extractos de soja pero no pudo ser determinada su
identidad fueron incluídos en la cantidad total de VOCs reportada.
Respuesta comportamental de insectos a los VOCs:
Bioensayos. Olfatómetro en "Y":
Tanto machos y
hembras adultos de P. guildinii
fueron colocados en el extremo basal de la “Y” del olfatómetro, donde se
encontraron frente a dos opciones: el blanco o el extracto de VOCs (muestreos R5
y R8).
Físicamente el olfatómetro
es un block de acrílico con una cavidad en forma de Y, aprisionado entre dos
placas planas de vidrio. El tronco del dispositivo mide 20 cm y cada brazo 17
cm (ver figura 7); tanto en el tronco como en cada brazo se trazaron
arbitrariamente líneas a la mitad de su longitud, para cuantificar la respuesta
del insecto.

Figura 7: Olfatómetro en Y donde se
realizaron los estudios comportamentales de preferencia de P. guildinii hacia
los VOCs de soja.
La fuente de olor se
generó absorbiendo 3 µl de la muestra (sea extracto o blanco de n-hexano) en un
papel de filtro (3 x 1 cm) doblado a la mitad a modo de carpa. Tanto los
papeles de filtro con el blanco como con el extracto fueron colocados en
cámaras cilíndricas de vidrio, las cuales tuvieron conexión con los respectivos
brazos del olfatómetro. El aire que arrastró los volátiles fue conducido por
caños de goma desde una bomba de diafragma (Aquarium Air Pump, LifeTech 9830),
pasando por un humidificador de agua destilada y un lecho de carbón activado
antes de llegar a las cámaras de volatilización. El flujo se reguló a 2.2
L/min, y se utilizó luz roja para emular condiciones nocturnas. Ello responde
al hecho de que en estudios preliminares de movilidad en olfatómetro en Y
(blanco contra blanco), los insectos fueron más móviles bajo luz roja que bajo
luz blanca.
La respuesta del
insecto se observó durante 15 minutos, registrándose la primera elección de
brazo, tiempo de residencia en cada brazo y número de veces que prefirió cada
uno de ellos. Cada individuo fue utilizado una única vez y entre ensayos sucesivos
se limpió el olfatómetro con etanol (70%), y se alternaron las cámaras de
volatilización.
La preferencia por
control o tratamiento se evaluó estadísticamente mediante el test de rangos
pareados de Wilcoxon para el número de entradas y tiempo de permanencia en cada
brazo. La primera elección de un brazo u otro se analizó por el test de Chi
cuadrado.
Análisis preliminar de los VOCs por GC-EAD:
Los ensayos
electrofisiológicos son claves para la Ecología Química. En los
electroantenogramas (EAG) se registra la diferencia de potencial entre los
extremos de la antena de un insecto como respuesta a un determinado estímulo.
Esta metodología puede ser acoplada a un cromatógrafo de gases (haciendo las
veces de detector conocido como EAD; ver figura 8), de manera de poder evaluar
la respuesta de la antena del insecto a los diferentes compuestos que eluyen de
la columna, simultáneamente a la detección de dichos compuestos por el detector
usual de GC.

Figura 8: cromatógrafo de gases acoplado
a detector de electroantenograma (GC-EAD). En la imagen de la izquierda se
puede observar en detalle la antena entre los electrodos.
Si algún compuesto
químico estimula dicha antena, ella emite un impulso eléctrico que es
amplificado y registrado por un software. De ésta manera se tiene una
comparación a tiempo real de los compuestos químicos eluídos y sus respectivas
respuestas de las antenas de los insectos.
Dado que las
cantidades de semioquímicos implicadas en las interacciones biológicas entre
organismos vivos son ínfimas (del orden de pg), tal detector es muy sensible.
Se pueden registrar respuestas electrofisiológicas en los cromatogramas en
donde no se puede visualizar ningún pico mediante los detectores usuales de GC.
Para llevar a cabo el
análisis se diseccionó la antena del insecto en su base, se la colocó entre dos
electrodos de plata y se la fijó a los mismos mediante un gel conductor. Este
es el encargado, en última instancia, de transmitir el impulso nervioso de la
antena a los electrodos. Los electrodos se conectaron a un amplificador, y éste
a un registrador en un PC, mediante
software HP Chem Station. Dado que la respuesta de las antenas no fue
consistente, se procedió a realizar un cambio en el sistema de detección,
utilizando como electrodos hilos de plata sumergidos en una solución llamada de
Ringer (solución de 0.755 % NaCl, 0.064 % KCl; 0.022 % CaCl2, 0.173
% MgCl2, 0.086% Na2HCO3 y 0.061 % Na3PO4)
que simula las condiciones osmóticas de la hemolinfa.
A los efectos de la
realización de éstos análisis, los insectos fueron colocados por 15 minutos en
una conservadora con escarchas de hielo, condiciones bajo las cuales los mismos
permanecen inmovilizados (entran en un estado de quiescencia), quedando aptos
para la disección de las antenas. Se procuró colocar la antena inmediatamente
después de la disección para formar parte del detector, y que la señal de la
misma permaneciera estable (visualizada en el registrador).
Para que la respuesta
detectada de la antena sea consistente, se pretende que la misma se repita en
al menos dos análisis consecutivos, lo cual no pudo ser verificado en ninguna
de las condiciones planteadas.
En éste
trabajo, debido a problemas de disponibilidad de insectos y de extractos, se
realizaron apenas 4 corridas de GCEAD con antenas de machos (dado que habían
mostrado una leve tendencia de atracción de VOCs de madurez en primera opción
de olfatómetro) y extractos de R8 (el más afectado por las chinches
en observaciones de campo).
Resultados



Resultados
Muestreo 1 (V4-V5):
En éste muestreo no
hubo diferencias apreciables entre el blanco y la muestra, detectándose dos
picos que no pudieron ser identificados por comparación con biblioteca de
espectros de masa.
Muestreo 2 (V8-V9):
En el mismo se
detectaron diferencias apreciables del extracto de VOCs de soja respecto del
blanco. Los compuestos identificados se ilustran en la figura 9.
La discriminación
según el tipo de compuestos identificados fue la siguiente:
Cetonas: 5-hepten2-ona–6-metil (GLV); geranil cetona.
Hidrocarburos monoterpenos: (E, β)
ocimeno.
Hidrocarburos sesquiterpenos: β- humuleno; α- zingibereno,.
Aldehídos: tetradecanal.
Ésteres: salicilato de metilo,
butanoato de 3-(Z)-hexen-1-ilo (GLV).
Éteres: epóxido de farneseno
(tentativo).

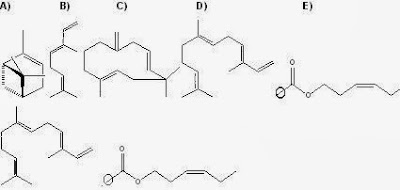
Figura 9:
VOCs
de soja identificados en el muestreo 2. A)
5-hepten2ona–6-metil, B) geranil
acetona, C) (E, β)-ocimeno, D) β-humuleno, E) α-zingibereno,
F) (E,E-α) farneseno G) tetradecanal H) salicilato de metilo I)
butanoato de 3-(Z)hexen-1-ilo.
Muestreo 3 (R2):
En éste muestreo
también se registraron diferencias apreciables respecto del blanco y de los
restantes muestreos.
La discriminación
según el tipo de compuestos identificados fue la siguiente:
Alcoholes: amil vinil carbinol (GLV).
Hidrocarburos monoterpenos: D-limoneno, (E,
β) ocimeno,
Hidrocarburos sesquiterpenos: α-copaeno, β-elemeno, β-humuleno; (E,E-α)
farneseno.
Ésteres: butanoato de 3-(Z)hexen-1-ilo (GLV).
Compuestos nitrogenados: farnesil
cianuro (tentativo).

Figura 10:
VOCs
de soja identificados en el muestreo 3. A)
amil vinil carbinol, B) limoneno, C) (E, β) ocimeno, D) α-copaeno E)
β-elemeno
F) β -humuleno, G) (E,E-α) farneseno, H)
butanoato de 3-(Z)-hexen-1-ilo.
Muestreo 4 (R5):
Si bien éste muestreo
fue en el que más se caracterizaron VOCs, la identificación se logró para pocos
de ellos. Existen diferencias apreciables tanto a nivel cuantitativo como
cualitativo respecto de los restantes muestreos.
La discriminación según
el tipo de compuestos identificados fue la siguiente:
Hidrocarburos aromáticos: tolueno (tentativo).
Cetonas: 3-hexanona (GLV) (tentativo).
Hidrocarburos monoterpenos: α-pineno, (E,
β)-ocimeno.
Hidrocarburos sesquiterpenos: β-humuleno; (E,E-α)
farneseno.
Ésteres: acetato de 3-(Z)hexen-1-ilo (GLV).
Aldehídos: 4-etilbenzaldehído
(tentativo).

Figura 11:
VOCs
de soja identificados en el muestreo 4. A) α-pineno,
B) (E, β)-ocimeno, C) β-humuleno, D)
(E,E-α)farneseno, E) acetato de 3-(Z)hexen-1-ilo.
Muestreo 5 (R8):
En éste muestreo,
correspondiente a la madurez fisiológica del cultivo, se obtuvieron muy pocos
VOCs. Aún así se pudieron identificar algunos no presentes en los restantes
muestreos como así también variación cuantitativa de compuestos comunes a los
otros muestreos.
La
discriminación según el tipo de compuestos identificados fue la siguiente:
Alcoholes: 3(Z)-hexen (1)ol (GLV).
Hidrocarburos
monoterpenos: (E, β)-ocimeno.
Ésteres: acetato de 3(Z)-hexen-1-ilo (GLV).
Aldehídos:
2-(E)hexenal (GLV).

Figura 12 VOCs
de soja identificados en el muestreo 5. A) 3(Z)-hexen-
(1)-ol , B) (E, β)ocimeno, C) acetato de 3(Z)-hexen-1-ilo, D) 2-(E)-hexenal.
Evolución de cantidades de los compuestos comunes
a muestreos:
Como pudo ser
apreciado anteriormente, hay ciertos compuestos comunes a los distintos
muestreos.
En éste apartado se
pretende representar las evoluciones individuales y colectivas de dichos VOCs
(figuras 13 y 14).
Se puede apreciar que
los compuestos (E,β)-ocimeno, (E,E-α)-farneseno y β-humuleno, siguen
aproximadamente la tendencia general de la evolución de las cantidades totales
de VOCs representada por el gráfico verde de la figura 13. En el mismo se puede
ver claramente un máximo cuantitativo de emisión en floración.
Sin embargo, los GLVs
butanoato de 3(Z)-hexen-1-ilo y
acetato de 3(Z)-hexen-1-ilo, presentan
el máximo de emisión en el estadio vegetativo avanzado y de llenado de vainas,
respectivamente.

Figura 13: Evolución
de los VOCs de soja (verde) y del (E,E-α)-farneseno (rojo) a lo largo
de la temporada de desarrollo del cultivo de soja. Cantidades equivalentes a ng
de tridecano.

Figura 14: Evolución de (E,β)-ocimeno
(A) y de β-humuleno (B) a lo largo del desarrollo fenológico de la soja. Las
cantidades se expresan en ng equivalentes de tridecano.
Respuesta comportamental de insectos a los VOCs:
Muestreo 4, R5.
A continuación se
presentan los resultados para machos y hembras para las tres variables medidas
en olfatómetro en Y.

Figura 15: Resultados
comportamentales en olfatómetro para machos (N=15) y hembras (N=8) de P.
guildinii en cuanto a número de entradas en cada compartimento (A), porcentaje de elección de primera opción (B) y tiempo de
permanencia en cada brazo (C) para los VOCs de R5. Las hembras se
visualizan en color claro y los machos en oscuro. Nota: para las hembras no se
realizó análisis estadístico debido al bajo número de individuos.
Muestreo 5, R8.
A continuación se
presentan los resultados para machos y hembras para las tres variables medidas
en olfatómetro en Y.

Figura 16: Resultados comportamentales
en olfactómetro para machos (N=15) y hembras (N=14) de P. guildinii en cuanto a
número de entradas en cada compartimento (A), porcentaje de elección de primera
opción (B) y tiempo de permanencia en cada brazo (C) para los VOCs de R8.
Las hembras se visualizan en color claro y los machos en oscuro.
Tratamiento estadístico e interpretación:
En el caso del
muestreo en R5 para los individuos machos, en las variables de
porcentaje de primera elección (Test Chi cuadrado; p> 0.4) y promedio de
número de entradas (Test de Wilcoxon; p>0.1) a cada brazo; hay una leve
tendencia aunque no significativa de atracción de los mismos hacia los VOCs de
soja. En el caso de la variable tiempo de residencia, tampoco hay diferencia
significativa entre ambos tratamientos, aunque la tendencia es a que la
permanencia en ambos brazos sea prácticamente la misma, según lo ilustra la
figura 15 C.
En el mismo muestreo
para las hembras, no se pueden hacer inferencias ninguna porque debido a
disponibilidad de insectos (en cantidad y estado fisiológico adecuado), no se
consiguió un número mínimo de individuos para lograr significancia estadística.
Aún, a pesar de lo antedicho, parece haber una tendencia de atracción hacia los
VOCs de soja en cuanto a porcentaje de elección de primera opción y tiempo de
permanencia en cada brazo.
Para el estadio R8
e individuos machos se observa disparidad en cuanto al valor de las distintas
variables, ya que hay una tendencia (nuevamente no significativa) a elegir
primera opción (Test Chi cuadrado; p> 0.1) con tratamiento de soja. Pero he
aquí que las otras dos variables, tanto promedio de número de entradas (Test de
Wilcoxon; p>0.1) y tiempo de residencia, sean favorables al blanco. En el
caso de la última variable, hay significancia estadística de repulsión a los VOCs
de soja (ejemplificado por el cuadro de error de la figura 16 C). Ello puede
interpretarse en el sentido de que el insecto sensa el ambiente del olfatómetro
y percibe un estímulo que lo repele y le hace huir de dicho brazo.
En el caso de las
hembras parece también haber una leve tendencia (no significativa) a la
repulsión de los VOCs, en las variables promedio de número de entradas (Test de
Wilcoxon; p>0.4) y primera opción (Test Chi cuadrado; p> 0.1).
Del estudio
bibliográfico de éste sistema y en la procura de las causas de éste
comportamiento inusual surgió que, el estándar interno usado (n-tridecano), es
uno de los principales compuesto defensivos de P. guildinii ante parasitoides, información ésta
que no se tenía al principio del trabajo. De ello se puede formular la
hipótesis de que el n-tridecano, aparte de ser un compuesto defensivo, pueda
actuar como feromona de alarma para los individuos de P. guildinii. Evidentemente para comprobar o refutar dicha
hipótesis se deberían hacer más bioensayos aumentando el número de individuos
testados.
Aún así, ello sirvió
de aprendizaje para los muestreos realizados en la temporada 2009-2010, los
cuales se hicieron con tetradecano como estándar interno. Los resultados de
dichos muestreos escapan a los objetivos y tiempo de duración de la tesis de
Licenciatura y no son presentados aquí.
Análisis de los VOCs por GC-EAD:
En las
corridas preliminares realizadas no pudo observarse ninguna respuesta
que indique atracción hacia alguno de los VOCs. Sin embargo, se sabe (González,
A.; comunicación personal), que las antenas de hemípteros no presentan padrones
de reacción tan claros como lepidópteros en EAD. Ello es cierto para feromonas,
y aún más para posibles kairomonas debido a que la relación planta-insecto no
es tan específica ni determinante para la sobrevivencia de la especie (P.
guildinii es polífaga) como la que surge de la emisión de aquellas.
Un aspecto
importante a destacar, entre
los factores bióticos que pueden influenciar los resultados aquí obtenidos, es
que al realizar el muestreo a campo el cultivo se encuentra sometido a un complejo
de plagas y no solamente a la plaga en estudio. Ejemplo de ello es que en la
temporada de muestreo se encontraron altas poblaciones de la arañuela (Tetranychus urticae, Acarina) y Anticarsia gemattalis (Lepidoptera).
Asimismo, no debe despreciarse la posibilidad de obtener VOCs de otras fuentes
durante los muestreos a campo, por ejemplo de la microfauna asociada a las
plantas, o del ambiente circundante (que incluye otras plantas tales como
malezas). A modo de ejemplo, compuestos como el 2-(E)-hexenal, α-pineno, α-zingibereno y (E,β)-ocimeno (informados aquí como VOCs
de soja), se han reportado
como emisiones de machos y hembras vírgenes de P. guildinii.
Perfil químico de VOCs:
En cuanto a la cantidad relativa de VOCs emitidos,
se pudo observar un aumento paulatino hasta el máximo en la floración, seguido
de un decrecimiento abrupto hasta la madurez fisiológica, como se puede
apreciar en la figura 13. Rostás et. al. reportó menor cantidad de VOCs con el
desarrollo fenológico de la soja , pero teniendo en cuenta
solamente los estadios V3 y R4 y no la floración.
En cuanto a la diversidad química detectada (medida
cómo la cantidad de compuestos diferentes), se puede observar un máximo en el
estadio de desarrollo de llenado de vainas (R5).
Según lo mostrado en las figuras 13 y 14, β-humuleno,
(E,E-α)-farneseno y β-E-ocimeno, parecen seguir (dentro de
ciertos límites de error experimental), la tendencia general de variación de
las cantidades relativas de VOCs mostrada en la figura 13. El β-E-ocimeno muestra un leve incremento hacia
el final de la temporada, lo que podría deberse a la emisión de P. guildinii, especie que alcanzó altas
poblaciones hacia el final de la temporada.
(E,E-α)-farneseno
aparece como el compuesto ampliamente mayoritario en los muestreos V8-V9
y R2, siendo mayor la proporción de este compuesto que la suma de
todos los restantes VOCs caracterizados en el muestreo 2 (56.5% del total de
VOCs) y el 46.2 % de los VOCs en el muestreo 3. En el muestreo correspondiente
a llenado de vainas (R5) es apenas el 11.5 % de los VOCs y no fue
detectado en la madurez fisiológica. Van de Boom halló dicho compuesto en una
proporción del 5% de los VOCs en soja vegetativa (de 5 semanas, posiblemente V3-V4),
donde es inducido por plantas sometidas a daño por T. urticae. Esto justificaría que
dicho compuesto no fuera detectado en el estadio vegetativo temprano en nuestro
estudio. Sin embargo, Rostás lo encontró como mayoritario en V3 ,
lo que puede reflejar propiamente el efecto de las condiciones ambientales y
las variedades de soja empleadas.
En el muestreo correspondiente al estado vegetativo
avanzado se pudo observar la presencia de salicilato de metilo (segundo
compuesto mayoritario, 5% del total de VOCs), un compuesto conocido por ser
elicitor de defensas químicas indirectas de la planta como consecuencia de la
herviboría de muchas especies, incluso de T.
urticae. Dicho compuesto ha sido reportado también como
atractivo a depredadores en interacciones tritróficas y se ha
planteado cómo un compuesto clave en la resistencia inducida contra ataques
fúngicos, virales o bacterianos.
El amil vinil carbinol (1-octen-3-ol) fue detectado
solamente en el muestreo correspondiente a floración. Sin embargo, Boué et.
al. determinaron que el mismo aumentaba a medida que el cultivo alcanza la
madurez por emisión específica de las semillas, lo que puede
deberse también a la variabilidad de éste tipo de compuestos.
El acetato de 3-(Z)-hexen-1-ilo,
un volátil de hoja verde (GLV), es el componente mayoritario del muestreo 4
(llenado de vainas), siendo el 26.9 % del total de los VOCs detectados, seguido
por el α-pineno con un 15.1%. Anteriormente Van Den Boom et al. reportaron al
primero de ellos cómo componente mayoritario de los VOCs inducidos por la
infestación de T. urticae en el estadio
vegetativo avanzado.
En el muestreo 5 (madurez fisiológica), el 2-(E)-hexenal (otro GLV) es el componente
mayoritario, alcanzando el 47.8 % de todos los VOCs. En éste muestreo se
detectan otros dos GLVs: 3-(Z)-hexen-(1)-ol
y acetato de 3-(Z)-hexen-1-ilo.
Si bien en todos los muestreos (salvo el primero) se
detectaron GLVs, resulta interesante que los mismos hayan sido detectados como
compuestos mayoritarios en los estados avanzados de madurez, donde no deberían
estar apreciablemente presentes, dado que las hojas verdes son escasas. Esto
puede deberse a la retención foliar de la soja, que hace que la misma llegue al
final de la temporada con hojas verdes, o a la emisión por parte de las
chinches que colonizaban las plantas hacia el final de la temporada.
El 2-(E)-hexenal
posee probada actividad antifúngica, siendo su
biosíntesis activada ante el ataque de herbívoros o patógenos. Mientras Boué
reporta una disminución del contenido de este aldehído en R7 y R8
respecto a R6 y a estadios vegetativos y de floración en soja no
atacada, en éste trabajo se encontró el patrón contrario, lo
que puede deberse a una respuesta de la planta inducida por el ataque de
plagas.
En el muestreo en R5 la cetona 3-hexanona
fue tentativamente identificada, y ha sido reportada anteriormente.
Dicho autor confirma la presencia de la misma en R6, R7 y
R8, con una disminución de la cantidad de la misma hasta alcanzar la
madurez, lo que explicaría que no se hubiera determinado en el presente trabajo
en R8. Incluso el mismo autor postula dicho compuesto como
quimioindicador de los estados de desarrollo R6 y R7.
Los compuestos nóveles respecto de reportes
anteriores fueron tetradecanal (V8-V9) α-copaeno, β-elemeno (R2) y tentativamente farnesil cianuro (V8-V9
y R2), tolueno y 4-etil benzaldehído.
La posible presencia de farnesil cianuro en el
estadio de floración no es sorprendente, ya que se han reportado varias oximas
y nitrilos dentro de los VOCs de soja, tanto de plantas sanas como dañadas.
Asimismo, dado que el (E,E-α)-farneseno es el compuesto mayoritario en ambos
estadios, no es de extrañar la presencia de vías metabólicas adicionales
activas para su biotransformación.
Estudios comportamentales:
En el muestreo en R5, P. guildinii, tanto en individuos machos
como hembras mostró una leve tendencia a
elegir como primera opción los extractos de soja, mientras que la diferencia en
el valor de las variables número de entradas a cada brazo y tiempo de
permanencia entre soja y blanco no es significativa.
En el muestreo en R8, por el contario, machos
y hembras muestran una leve tendencia a entrar más veces al brazo del blanco
que al brazo de los VOCs de soja. En cuanto al tiempo de residencia, hay una
tendencia significativa de repulsión de machos hacia el brazo de los VOCs. Mientras
tanto, en la elección de primera opción se obtuvieron resultados contrapuestos
entre machos y hembras, los machos con tendencia de atracción hacia los VOCs de
soja y las hembras de repulsión a los mismos.
Lo antedicho permite apreciar lo complejo que puede
ser el estudio del comportamiento animal. Aún así en éste caso puede haber
incidencia directa del estándar interno en dicho comportamiento, ya que por
falta de información se incurrió a uno de los principales compuestos defensivos
secretados por las chinches.
Conclusiones y Perspectivas
En éste trabajo se identificaron VOCs de soja
reportados anteriormente y se reporta la existencia de algunos volátiles
nuevos.
La evaluación comportamental hacia los VOCs, parece
ser poco clara con resultados contrapuestos para R5 y R8,
aunque puede haberse debido a errores de la metodología utilizada. Los
estudios electroantenográficos hasta el momento no han arrojado ningún
resultado concluyente.
Aunque
no se ha obtenido evidencia comportamental y electrofisiológica, se seguirá
investigando sobre las bases químicas de la comunicación interespecífica entre P.
guildinii y G. max. Una aliciente importante para éste trabajo es el
hecho de que a campo se ha observado (Castiglioni, E.; comunicación personal)
un rápido aumento de la población de la plaga en un corto período de tiempo
posterior a la floración. Las bases de éste comportamiento pueden subyacer en
la percepción por parte de los insectos de un estímulo químico proveniente de
la planta (kairomona) que ha sido adaptativamente seleccionado para la
ubicación de la fuente de alimentación.



Comentarios
Publicar un comentario